


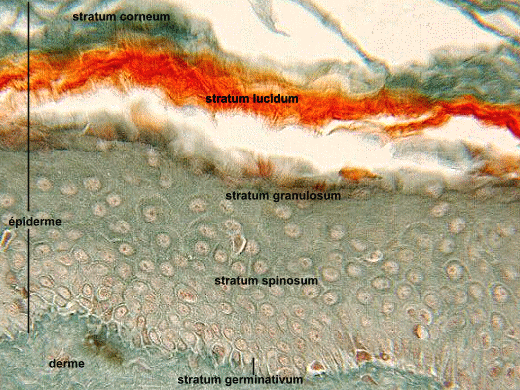
L'épiderme
L’épiderme est la région la plus superficielle. L’épiderme est un épithélium de revêtement, stratifié, pavimenteux et orthokératosique. C’est un épithélium non vascularisé. Il est constitué de 4 types cellulaires :
I.1.1 Les kératinocytes :
Les kératinocytes représentent 80% de l’ensemble des cellules de l’épiderme. Ce sont eux qui en migrant, donnent à l’épiderme ses caractéristiques morphologiques : stratification en plusieurs couches et cellules superficielles pavimenteuses et anuclées.
D’origine ectoblastique, ce sont les principales cellules de l’épiderme et assurent 3 grandes fonctions :
- La cohésion de l’épiderme en rapport avec le cytosquelette et les systèmes de jonction des kératinocytes entre eux.
- Une fonction de barrière entre les milieux intérieur et extérieur.
- La protection contre les radiations lumineuses en rapport avec les mélanosomes.
Les kératinocytes se répartissent en 4 couches qui sont, de la profondeur à la superficie, la couche basale, la couche spineuse, la couche granuleuse et la couche cornée (compacte puis desquamante). L’épaisseur des couches est variable suivant l’âge de l’individu et les régions du corps, elle est maximum au niveau de la peau épaisse.
§ La couche basale :
Elle est formée de l’ensemble des kératinocytes directement en contact avec la jonction dermo-épidermique. Les kératinocytes forment une seule assise de cellules cylindriques et comprennent 3 populations :
- Les cellules souches de l’épiderme, particulièrement abondantes au niveau des crêtes épidermiques interpapillaires.
- Les cellules amplificatrices se divisant avant d’entrer dans le compartiment de différenciation.
- Les cellules post-mitotiques qui restent en position basale.
Les mélanosomes sont phagocytés par les kératinocytes basaux. Ils disparaissent du cytoplasme des kératinocytes dans les couches supra-basales : cette disparition est rapide dans les peaux claires, lente dans les peaux foncées ou bronzées.
Les hémidesmosomes et les desmosomes sont les systèmes de jonction sur lesquels s’accrochent les tonofilaments : les hémidesmosomes accrochent les kératinocytes à la matrice cellulaire alors que les desmosomes accrochent les kératinocytes entre eux (peu nombreux dans la couche basale).
§ La couche spineuse :
Les kératinocytes qui ont quitté la couche basale deviennent polygonaux tandis que leur noyau s’arrondit et leur cytoplasme devient plus foncé : la rétraction du cytoplasme due à des artéfacts de préparation en microscopie optique donne un aspect épineux à ces cellules, d’où le nom de la couche. C’est la couche qui contient le plus de desmosomes.
§ La couche granuleuse :
Le cytoplasme et le noyau des kératinocytes s’aplatissent. C’est l’apparition de granulations basophiles dans le cytoplasme des kératinocytes qui définit cette couche. Ce sont les grains de kératohyaline et les kératinosomes qui sont carctéristiques et spécifiques des kératinocytes de la couche granuleuse de l’épiderme. Ils disparaissent dans la couche cornée. Les grains de kératohyaline correspondent aux grains basophiles. Les kératinosomes (trop petits pour être vus en microscopie optique) migrent progressivement de la région périnucléaire à proximité de l’appareil de Golgi vers la membrane cytoplasmique avec laquelle ils fusionnent déversant alors leur contenu dans l’espace extracellulaire. Ainsi les kératinosomes sont à l’origine du cément entre les cornéocytes.
§ La couche cornée :
Finalement, les kératinocytes perdent brutalement leur noyau, ils deviennent des cornéocytes qui constituent la couche cornée, compacte en profondeur au contact de la granuleuse, desquamante en superficie. Finalement, le cytoplasme des cornéocytes devient floconneux en même temps que se lysent le cément intercellulaire et les cornéodesmosomes, ce qui aboutit à la desquamantation des cornéocytes les plus superficiels.
La couche cornée est donc un ensemble de cellules « mortes » mais fonctionnelles réunies entre elles par un cément, l’ensemble donnant à l’épiderme sa fonction de barrière.
I.1.2 Les éléments de jonction :
- Les molécules des tonofilaments qui sont des filaments intermédiaires, sont comme dans tous les épithéliums, des kératines presque toujours associées en paires. Il s’agit de la paire K5-K14 et K15 dans la couche basale et K1-K10 et K2-K11 spécifiques de l’épiderme dans les couches supra-basales. Ces tonofilaments jouent un rôle important pour les hémidesmosomes et les desmosomes.

Kératinocyte de la couche basale
de l'épiderme en microscopie électronique
1 = tonofilament
2 = desmosome
3 = hémidesmosome
4 = mélanosome
- Les molécules des desmosomes se répartissent principalement en molécules transmembranaires et en molécules de plaques :
- Les principales molécules transmembranaires sont les desmogléines qui appartiennent à la famille des cadhérines desmosomales, établissant entre elles des liaisons homophiliques.
- Les principales molécules de plaques sont les desmoplakines, l’envoplakine, la périplakine la plakoglobine et les plakophillines.
- Un troisième type de molécules a été mis en évidence. Elle apparaît dans la couche spineuse, à la partie supérieure.
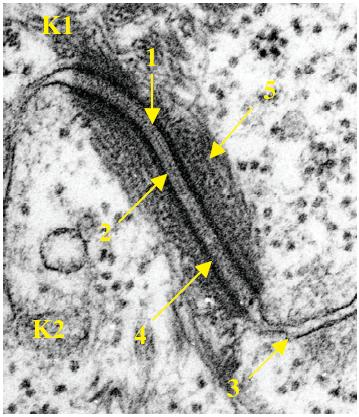
Desmosome en microscopie électronique:
1 = ligne dense extracellulaire
2 = desmogléie
3 = membrane cytoplasmique
4 = plaque
5 = tonofilaments
I.1.3 Les mélanocytes :
Les mélanocytes constituent la 2ème grande population cellulaire de l’épiderme. Ils proviennent des crêtes neurales et ne colonisent que secondairement l’épiderme où, à terme, ils sont exclusivement situés dans la couche basale de l’épiderme. Leur fonction est la synthèse des mélanines : phéomélanines et eumélanines, dans des organites spécialisés, les mélanosomes qui sont ensuite transférés aux kératinocytes. Les mélanines ont 2 fonctions : elles donnent à la peau sa « couleur », les phéomélanines étant des pigments jaunes-rouges et les eumélanines, des pigments brun-noirs ; la pigmentation constitutive s’oppose à la pigmentation « facultative » communément appelée bronzage qui apparaît après l’irradiation des UV. Les eumélanines ont un rôle photoprotecteur. En revanche, sous l’action des radiations lumineuses, les phéomélanines sont cancérigènes. La répartition entre les eumélanines et les phéomélanines varie selon les individus et conditionne leur phototype cutané. Par convention, en fonction de la couleur constitutive de la peau et de ses capacités à développer une pigmentation sous l’effet des rayons UV. Les UV n’agissent pas directement sur les mélanocytes mais agissent indirectement par l’intermédiaire des kératinocytes qui sécrètent en particulier de l’α-MSH, capable de se fixer sur le recepteur MC1-R des mélanocytes. On distingue 6 phénotypes cutanés

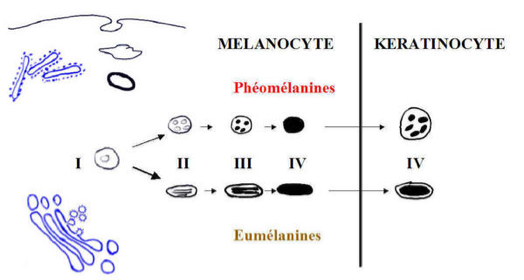
La synthèse des mélanines :
La synthèse de toutes les mélanines commencent par l’hydroxylation de la tyrosine en DOPA sous l’action d’une tyrosinase, puis l’oxydation de la DOPA en dopaquinone sous l’action de cette même enzyme. Ainsi la DOPA réaction est une réaction histochimique spécifique des mélanocytes. La poursuite de la synthèse se fait vers la voie des phéomélanines et/ou la voie des eumélanines. La dopaquinone entre dans la voie des pheomélanines si elle rencontre une grande quantité de cystéine, sinon elle s’oriente dans la voie des eumélanines où une enzyme de la même famille que la tyrosinase, la TRP2 intervient.
La maturation des mélanosomes :
Il y a 4 stades de maturation des mélanosomes : les stades I et II correspondent à la synthèse de l’organite, qui contient la tyrosinase non active. Le stade III à la synthèse des mélanines après activation de la tyrosinase. Le stade IV à un mélanosome complètement mélanisé où la tyrosinase n’est plus active, ce qui explique la négativité de la DOPA réaction dans les kératinocytes.

I.1.4 Les cellules de Langerhans
Les cellules de Langerhans sont la 3ème population cellulaire de l’épiderme (3 à 8% des cellules épidermiques). Elles appartiennent au groupe des cellules dendritiques présentatrices d’antigènes aux lymphocytes T. Les cellules de Langerhans sont d’abord produites au niveau des organes hématopoïétiques. Elles vont ensuite migrer vers l’épiderme, s’y installer.
En microscopie optique, les cellules de Langerhans apparaissent comme des cellules claires, à noyau encoché, situées le plus souvent au niveau de la couche granuleuse de l’épiderme.
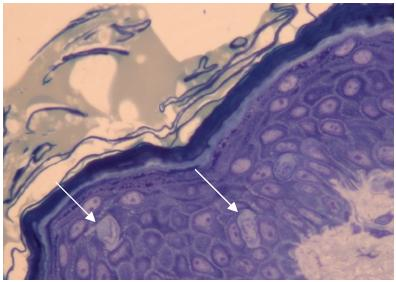
Cellules de Langerhans en microscopie optique - mise en évidence: bleu de toluidine -
I.1.5 Les cellules de Merkel
Les cellules de Merkel constituent la 4ème population cellulaire de l’épiderme. Ce sont des cellules neuro-épithéliales, dérivant des cellules souches de l’épiderme fœtal, qui ont pour fonctions celles de mécanorécepteurs à adaptation lente de type 1 et/ou des fonctions inductives et trophiques sur les terminaisons nerveuses périphériques et les annexes cutanées.
Les cellules de Merkel ne sont pas visibles en microscopie optique standart.
En microscopie électronique, ce sont des cellules ovalaires, à grand axe souvent parallèle à la jonction dermo-épidermique, à noyau dense, contourné ou indenté.

I.1.6 Pathologies de l’épiderme
Par convention, on appelle :
- hyperkératose, un épaississement de la couche cornée, elle est orthokératosique lorsque les cornéocytes ne présentent pas de noyau et parakératosique lorsque ces derniers sont présents.
- agranulose : l’absence de couche spineuse.
- hyperacanthose : l’épaississement de la couche spineuse.
- hyperpapillomatose : l’accentuation des ondulations de la jonction dermo-épidermique avec allongement des papilles dermiques et des crêtes épidermiques.
- acanthylose : séparation des kératinocytes entre eux par rupture des desmosomes.
Ces termes histologiques sont utilisés dans les comptes tendus d’anatomopathologie d’une biopsie cutanée.
- Les carcinomes baso-cellulaires :
Les carcinomes baso-cellulaires sont des tumeurs cutanées en rapport avec une prolifération des kératinocytes de la couche basale de l’épiderme.
- Les dermatoses bulleuses intra-épidermiques
Les altérations des structures assurant la cohésion de l’épiderme aboutissent en règle générale à des dermatoses bulleuses intra-épidermiques, d’origine génétique ou auto-immune.
- les dermatoses bulleuses intra-épidermiques héréditaires : sont en rapport avec des anomalies génétiques des cytokératines dont les plus fréquentes sont des lyses bulleuses héréditaires (EBH) simples par mutation sur les gènes des kératines K5 ou K14.
La « maladie des conscrits » est une EBH simple à transmission dominante très fréquente se traduisant par la formation inhabituelles « d’ampoules » au niveau des pieds au décours de marches prolongées. Elle est en rapport avec des mutations sur les gènes des kératines K5 ou K14.
Les maladies touchant les kératines K1, K2, K6, K9, K10, K16 et K17 sont plus rares.
-Les dermatoses bulleuses intra-épidermiques, dites auto-immunes : sont en rapport avec la fixation d’auto anticorps dirigés contre les molécules des desmosomes. Ce sont les pemphigus. Le pemphigus vulgaire, qui est relativement fréquent en France, touche la peau et toutes les muqueuses malpighiennes, est secondaire à la formation d’anticorps dirigés contre la desmogléine (Dsg3).
Plus récemment, des anomalies des molécules de la cohésion épidermique ont aussi été démontrées dans des pathologies hyperkératosiques :
- la dysplasie ectodermique avec fragilité cutanée, par mutation sur le gène de la plakophiline 1.
- les kératodermies palmoplantaires striées, par mutation sur le gène de la desmoplakine.
- la maladie de Carjaval Huerta par mutation sur le gène de la desmoplakine.
- la maladie de Naxos par mutation sur le gène de la plakoglogine.
Les albinismes sont la conséquence d’anomalie dans la mélanogénèse.





